The endometrium-specific proteome
|
The endometrial mucosa lines the inner wall of the uterus and prevents adhesion between the opposed walls to maintain the patency of the uterine cavity. The endometrium in fertile women undergoes hormone-driven cyclic regeneration, coarsely divided into a menstrual, a proliferative and a secretory phase. The transcriptome analysis shows that 72% of all human proteins (n=19628) are expressed in the endometrium and 116 of these genes show an elevated expression in endometrium compared to other tissue types.
An analysis of the genes with elevated expression in the endometrium shows differences in expression depending on the menstrual cycle.
|
- 4 endometrium enriched genes
- 116 genes defined as elevated in the endometrium
- Most of the elevated genes encode secreted proteins
- Most group enriched genes share expression with smooth muscle
|
Figure 1. The distribution of all genes across the five categories based on transcript abundance in endometrium as well as in all other tissues.
116 genes show some level of elevated expression in the endometrium compared to other tissues. The three categories of genes with elevated expression in endometrium compared to other organs are shown in Table 1.
Table 1. The genes with elevated expression in endometrium
Category |
Number of genes |
Description |
| Tissue enriched |
4 |
At least five-fold higher mRNA levels in a particular tissue as compared to all other tissues |
| Group enriched |
26 |
At least five-fold higher mRNA levels in a group of 2-7 tissues |
| Tissue enhanced |
86 |
At least five-fold higher mRNA levels in a particular tissue as compared to average levels in all tissues |
| Total |
116 |
Total number of elevated genes in endometrium |
Table 2. The 4 genes with the highest level of enriched expression in endometrium. "Predicted localization" shows the classification of each gene into three main classes: Secreted, Membrane, and Intracellular, where the latter consists of genes without any predicted membrane and secreted features. "mRNA (tissue)" shows the transcript level as TPM values, TS-score (Tissue Specificity score) corresponds to the score calculated as the fold change to the second highest tissue.
Gene |
Description |
Predicted localization |
mRNA (tissue) |
TS-score |
| MMP26 |
matrix metallopeptidase 26 |
Secreted |
23.8 |
64 |
| MMP10 |
matrix metallopeptidase 10 |
Secreted |
443.1 |
39 |
| PAEP |
progestagen-associated endometrial protein |
Intracellular,Secreted |
437.4 |
7 |
| IL17C |
interleukin 17C |
Secreted |
3.5 |
6 |
Some of the proteins predicted to be membrane-spanning are intracellular, e.g. in the Golgi or mitochondrial membranes, and some of the proteins predicted to be secreted can potentially be retained in a compartment belonging to the secretory pathway, such as the ER, or remain attached to the outer face of the cell membrane by a GPI anchor.
The endometrium transcriptome
An analysis of the expression levels of each gene makes it possible to calculate the relative mRNA pool for each of the categories. The analysis shows that 87% of the mRNA molecules derived from endometrium correspond to housekeeping genes and only 1% of the mRNA pool corresponds to genes categorized as endometrium enriched, group enriched, or endometrium enhanced. Thus, most of the transcriptional activity in the endometrium relates to proteins with presumed housekeeping functions as they are found in all tissues and cells analyzed.
Protein expression of genes elevated in the endometrium
In-depth analysis of the elevated genes in endometrium using antibody-based protein profiling allowed us to create a map of where these proteins are expressed within the endometrium, including glandular cells, stroma cells and with respect to differences dependent on the menstrual cycle.
Proteins expressed during the proliferative phase
Proteins expressed during the proliferative phase include PGR, the progesterone receptor which plays a central role in reproductive events, expressed in both glandular and stroma cells, and TRH, a thyrotropin-releasing hormone involved in the regulation and release of thyroid-stimulating hormone as well as prolactin, expressed in glandular cells.
Proteins specifically expressed during the secretory phase
Proteins expressed during the secretory phase include PAEP, a glycoprotein with essential roles in regulating a uterine environment suitable for pregnancy, expressed in glandular cells and C1QTNF6, a complement C1q tumor necrosis factor-related protein with unknown function, expressed in stroma cells.
Proteins specifically expressed during both secretory and proliferative phase
Proteins expressed during both the proliferative and secretory phase include HOXA11, a transcription factor involved in the regulation of uterine development and required for female fertility, expressed in stroma cells, TMEM200B a transmembrane protein with unknown function, expressed in glandular cells and SFRP4, a modulator of Wnt signaling with a role in regulating cell growth and differentiation in specific cell types, expressed both in stroma and glandular cells.
Genes shared between the endometrium and other tissues
There are 26 group enriched genes expressed in the endometrium. Group enriched genes are defined as genes showing a 5-fold higher average level of mRNA expression in a group of 2-7 tissues, including endometrium, compared to all other tissues.
In order to illustrate the relation of the endometrium tissue to other tissue types, a network plot was generated, displaying the number of commonly expressed genes between different tissue types.
Figure 2. An interactive network plot of the endometrium enriched and group enriched genes connected to their respective enriched tissues (grey circles). Red nodes represent the number of endometrium enriched genes and orange nodes represent the number of genes that are group enriched. The sizes of the red and orange nodes are related to the number of genes displayed within the node. Each node is clickable and results in a list of all enriched genes connected to the highlighted edges. The network is limited to group enriched genes in combinations of up to 3 tissues, but the resulting lists show the complete set of group enriched genes in the particular tissue.
The endometrium shares a few genes with most other tissue types but has most shared genes with smooth muscle (12) and other female tissues including placenta (5 genes) and fallopian tube (7 genes).
Endometrium shows a close relation to other female tissues, e.g. ESR1, the estrogen receptor
alpha, which is expressed in various cell types in female tissues. Another example is
EMX2, which shows group enriched expression in eight different tissues, including endometrium,
fallopian tube and cervix.
MSX1 is a homeobox transcription factor known to be involved in craniofacial development and odontogenesis. MSX1 is also implicated in the development of the nail bed mesenchyme. Our analysis shows that MSX1 is group enriched in the cervical and endometrial glands.
Another example of a gene shared between endometrium and other tissues is GNLY, a secreted protein located in cytotoxic granules of T-lymphocytes. GNLY shows elevated expression in endometrium and is also highly expressed in bone marrow and spleen. Interestingly, this is not the case for CD8 (CD8A), a common marker of cytotoxic T-lymphocytes.
Endometrium function
The endometrium is the inner mucous membrane of the uterus and consists of endometrial glands and endometrial stroma. It functions as a lining for the uterus, preventing adhesions between the opposed walls of the myometrium, thereby maintaining the patency of the uterine cavity.
Menstrual cycle
The endometrial lining undergoes cyclic regeneration. The menstrual cycle in the uterine can be divided into menstruation, proliferative phase, and secretory phase and is controlled by the endocrine system.
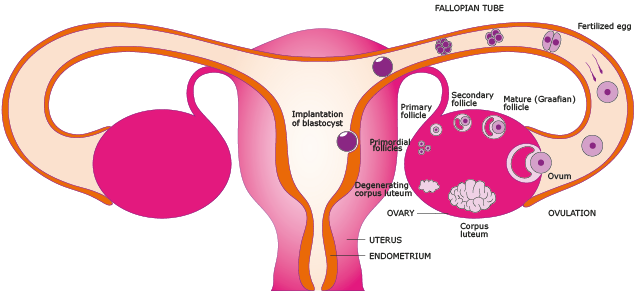
During the proliferative phase, as a response to increasing amounts of estrogen, the lining of the endometrium grows to a thick, blood vessel-rich, glandular tissue layer. Under complex hormonal inter-play follicles in the ovary start to develop. As they mature, the ovarian follicles secrete increasing amounts of estrogens which initiate the formation of a new layer of endometrium in the uterus, histologically identified as the proliferative endometrium. The non-dominant ovarian follicles atrophy and die whereas the dominant follicle releases an ovum (egg).
During the secretory phase pituitary hormones cause the remaining parts of the dominant follicle to transform into the corpus luteum, which produces progesterone. After ovulation and a rise in progesterone levels by the corpus luteum the endometrium changes to prepare for implantation of the blastocyst (fertilized egg). If implantation does not occur within approximately two weeks the levels of progesterone and estrogen drops causing the uterus to shed its lining and egg in menstruation.
During pregnancy, the glands and blood vessels in the endometrium continue to increase in size and number. Under the influence of progesterone, in a process called decidualization, the vascular spaces fuse and become interconnected, forming the maternal part of the placenta, the decidua, which supplies oxygen and nutrition to the fetus.
Figure 3. The ovaries, fallopian tubes and uterus. The menstrual cycle can be described both by the ovarian cycle and the uterine cycle which consists of menstruation, proliferative phase, and secretory phase. During the proliferative phase the lining of the uterus grow, or proliferate, and follicles in the ovary start to mature. During ovulation the dominant follicle releases an ovum (egg) via the fallopian tube. After ovulation and during the secretory phase the remaining parts of the follicle transforms into the corpus luteum which produces hormones that support the early pregnancy or if the ovum is not fertilized will atrophy leading to falling levels of hormones and the beginning of menstruation and a new cycle. If fertilized the ovum develops into a blastocyst and implantation in the uterus is made possible through changes in both the blastocyst and endometrial wall. Implantation allows for the next step in embryogenesis which includes formation of the placenta.
Endometrium histology
The inactive, prepubertal endometrium shows a cuboidal to low columnar epithelium that lines the surface and the underlying glands. The appearance greatly resembles the inactive endometrium seen in postmenopausal women, as both prepubertal and postmenopausal endometrium does not exhibit any proliferative or secretory changes that are hormone dependent. The endometrium in the reproductive female may be considered to comprise of a deeper basal layer and a superficial functional layer. The functional layer is subdivided into two strata - the compactum (towards the surface) and the spongiosum (towards the basalis).
The histology of human endometrium including detailed images and information can be viewed in the Protein Atlas Histology Dictionary.
Background
Here, the protein-coding genes expressed in the endometrium are described and characterized, together with examples of immunohistochemically stained tissue sections that visualize protein expression patterns of proteins that correspond to genes with elevated expression in the endometrium.
Transcript profiling and RNA-data analyses based on normal human tissues have been described previously (Fagerberg et al., 2013). Analyses of mRNA expression including over 99% of all human protein-coding genes was performed using deep RNA sequencing of 172 individual samples corresponding to 37 different human normal tissue types. RNA sequencing results of 5 fresh frozen tissues representing normal endometrium was compared to 167 other tissue samples corresponding to 36 tissue types, in order to determine genes with elevated expression in endometrium. A tissue-specific score, defined as the ratio between mRNA levels in endometrium compared to the mRNA levels in all other tissues, was used to divide the genes into different categories of expression.
These categories include: genes with elevated expression in endometrium, genes expressed in all tissues, genes with a mixed expression pattern, genes not expressed in endometrium, and genes not expressed in any tissue. Genes with elevated expression in endometrium were further sub-categorized as i) genes with enriched expression in endometrium, ii) genes with group enriched expression including endometrium and iii) genes with enhanced expression in endometrium.
Human tissue samples used for protein and mRNA expression analyses were collected and handled in accordance with Swedish laws and regulation and obtained from the Department of Pathology, Uppsala University Hospital, Uppsala, Sweden as part of the sample collection governed by the Uppsala Biobank. All human tissue samples used in the present study were anonymized in accordance with approval and advisory report from the Uppsala Ethical Review Board.
Relevant links and publications
Uhlén M et al, 2015. Tissue-based map of the human proteome. Science
PubMed: 25613900 DOI: 10.1126/science.1260419 Yu NY et al, 2015. Complementing tissue characterization by integrating transcriptome profiling from the Human Protein Atlas and from the FANTOM5 consortium. Nucleic Acids Res.
PubMed: 26117540 DOI: 10.1093/nar/gkv608 Fagerberg L et al, 2014. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol Cell Proteomics.
PubMed: 24309898 DOI: 10.1074/mcp.M113.035600 Zieba A et al, 2015. The Human Endometrium-Specific Proteome Defined by Transcriptomics and Antibody-Based Profiling. OMICS.
PubMed: 26488136 DOI: 10.1089/omi.2015.0115 Histology dictionary - the endometrium
|